
Sediment records of environmental change in Antarctica
Dr Brendan Keely
University of York
Chairman of the Environmental Chemistry Group
ECG bulletin January 2008
University of York
Chairman of the Environmental Chemistry Group
ECG bulletin January 2008
Dr Brendan Keely from the University of York, and the current Chairman of the Environmental Chemistry Group, explains how the pattern of substitution of porphyrin pigment derivatives found in sediments extracted from Antarctic lakes is a sensitive marker of past climates and environmental change.
Introduction
Climate variability. In recent years, scientists, the public and policy makers have become keenly aware of the far-reaching consequences that climate change could have for society in the near future. Notably, parts of the Antarctic are already experiencing rapid warming events that have occurred at a greater rate than in other parts of the Earth [1].
Since the ice cover in the region represents over 70% of the world’s freshwater, and some is close to the melting point, rising temperatures have the potential to cause the release of considerable volumes of freshwater to the oceans. The consequent change in the balance of energy exchange between the ice, ocean and atmosphere would have significant impact on the world’s climate. Records of past natural climate variability hold the key to understanding how temperature changes affecting Antarctica would influence other regions of the Earth.
Influence of the southern hemisphere on climate. The Quaternary (the last 1.7 million years) was marked by several glacial/interglacial events. Evidence suggests that the last two deglaciation events in the southern hemisphere triggered major changes in climate, including abrupt climate warming, in the northern hemisphere [2, 3].
The Holocene (the last 11,500 years) was marked by many, often rapid, temperature excursions that were of sufficient magnitude to affect human populations through drought, crop failure, floods and extreme weather events. In order to recover temporally detailed and well-resolved records of environmental change, such as changes in relative sea-level during the Holocene and beyond, it is necessary to have highly sensitive and universally applicable methods that can be applied to sediments.
Lake sediments as records of environmental change. Sediments have the potential to record and store long-term records of environmental conditions. Lake sediments are particularly valuable since they often contain undisturbed records and the origins of the sediments are constrained by the lake morphology and location. Lake sediments contain organic residues originating from the various natural product compounds produced by the organisms that inhabited the water column at the time the sediment was deposited. Sedimentary organic compounds that show structural relationships to the natural product from which they originated can be used as biological markers to provide information on the composition of the biological community.
Accordingly, depth profiles of biological markers through cores of lake bottom sediments reveal changes in the populations of aquatic organisms over time. Such changes in population are driven mainly in response to changes in environmental conditions.
Chlorophyll pigments as biological markers
Derivatives of chlorophyll pigments are particularly useful biological markers [4]. Because they originate exclusively from the primary producer community, the pigments reflect the base of the food chain and hence the major groups of organisms that inhabited the lake waters at any point in time. Based on the nature of the tetrapyrrole macrocycle, several distinct types of chlorophyll pigment can be recognised in extant organisms, including pigments unique to eukaryotes (e.g. chlorophylls a and b; Figure 1) and others that exist only in photosynthetic bacteria (e.g. bacteriochlorophylls c and d; Figure 2).
are driven mainly in response to changes in environmental conditions.
Introduction
Climate variability. In recent years, scientists, the public and policy makers have become keenly aware of the far-reaching consequences that climate change could have for society in the near future. Notably, parts of the Antarctic are already experiencing rapid warming events that have occurred at a greater rate than in other parts of the Earth [1].
Since the ice cover in the region represents over 70% of the world’s freshwater, and some is close to the melting point, rising temperatures have the potential to cause the release of considerable volumes of freshwater to the oceans. The consequent change in the balance of energy exchange between the ice, ocean and atmosphere would have significant impact on the world’s climate. Records of past natural climate variability hold the key to understanding how temperature changes affecting Antarctica would influence other regions of the Earth.
Influence of the southern hemisphere on climate. The Quaternary (the last 1.7 million years) was marked by several glacial/interglacial events. Evidence suggests that the last two deglaciation events in the southern hemisphere triggered major changes in climate, including abrupt climate warming, in the northern hemisphere [2, 3].
The Holocene (the last 11,500 years) was marked by many, often rapid, temperature excursions that were of sufficient magnitude to affect human populations through drought, crop failure, floods and extreme weather events. In order to recover temporally detailed and well-resolved records of environmental change, such as changes in relative sea-level during the Holocene and beyond, it is necessary to have highly sensitive and universally applicable methods that can be applied to sediments.
Lake sediments as records of environmental change. Sediments have the potential to record and store long-term records of environmental conditions. Lake sediments are particularly valuable since they often contain undisturbed records and the origins of the sediments are constrained by the lake morphology and location. Lake sediments contain organic residues originating from the various natural product compounds produced by the organisms that inhabited the water column at the time the sediment was deposited. Sedimentary organic compounds that show structural relationships to the natural product from which they originated can be used as biological markers to provide information on the composition of the biological community.
Accordingly, depth profiles of biological markers through cores of lake bottom sediments reveal changes in the populations of aquatic organisms over time. Such changes in population are driven mainly in response to changes in environmental conditions.
Chlorophyll pigments as biological markers
Derivatives of chlorophyll pigments are particularly useful biological markers [4]. Because they originate exclusively from the primary producer community, the pigments reflect the base of the food chain and hence the major groups of organisms that inhabited the lake waters at any point in time. Based on the nature of the tetrapyrrole macrocycle, several distinct types of chlorophyll pigment can be recognised in extant organisms, including pigments unique to eukaryotes (e.g. chlorophylls a and b; Figure 1) and others that exist only in photosynthetic bacteria (e.g. bacteriochlorophylls c and d; Figure 2).
are driven mainly in response to changes in environmental conditions.
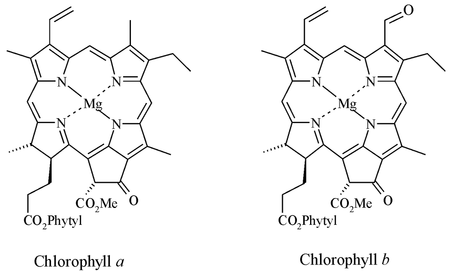
Figure 1: The structure of chlorophyll a and chlorophyll b
|
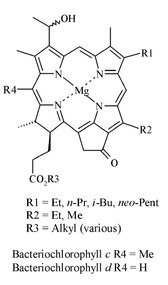
Figure 2: The structure of bacteriochlorophyll c and bacteriochlorophyll d
|

Figure 3: Structure of a chlorin steryl ester (a sterol ester-linked via the 3 position to pyrophaeophorbide a)
|
Some of the bacteriochlorophylls also exhibit variations in the nature of peripheral substituents (R1-3 in Figure 2). In some cases in excess of 50 individual compounds are possible. Laboratory and field studies indicate that at least some of the structural variations result in response to changes in environmental conditions, including the intensity and spectral profile of the photosynthetically active radiation available to the organisms [5, 6]. Furthermore, various chemical modifications to the pigments occur during senescence (cellular disruption associated with death), during grazing by zooplankton, and as the organic debris in which they are contained sinks through the water column.
|
Common processes involve replacement of Mg by 2H to form phaeophytins, further loss of CO2Me to form pyrophaeophytins and hydrolysis of the phytyl ester to form phaeophorbides. Particular chemical structures formed during these transformations also include steryl chlorin esters (SCEs; Figure 3) and oxidation products (e.g. purpurins; Figure 4) [4].
Steryl chlorin esters are a class of chlorophyll transformation product in which the sterol moiety is ester-linked to a chlorin carboxylic acid, most often pyrophaeophorbide a (Figure 3). SCEs essentially arise from the grazing of phytoplankton by zooplankton, and occur widely in marine and lacustrine (lake) sediments. The sterol portion of the SCEs derives from either the algal community or a zooplankton grazer, and is introduced into the pigment structure via transesterification. |
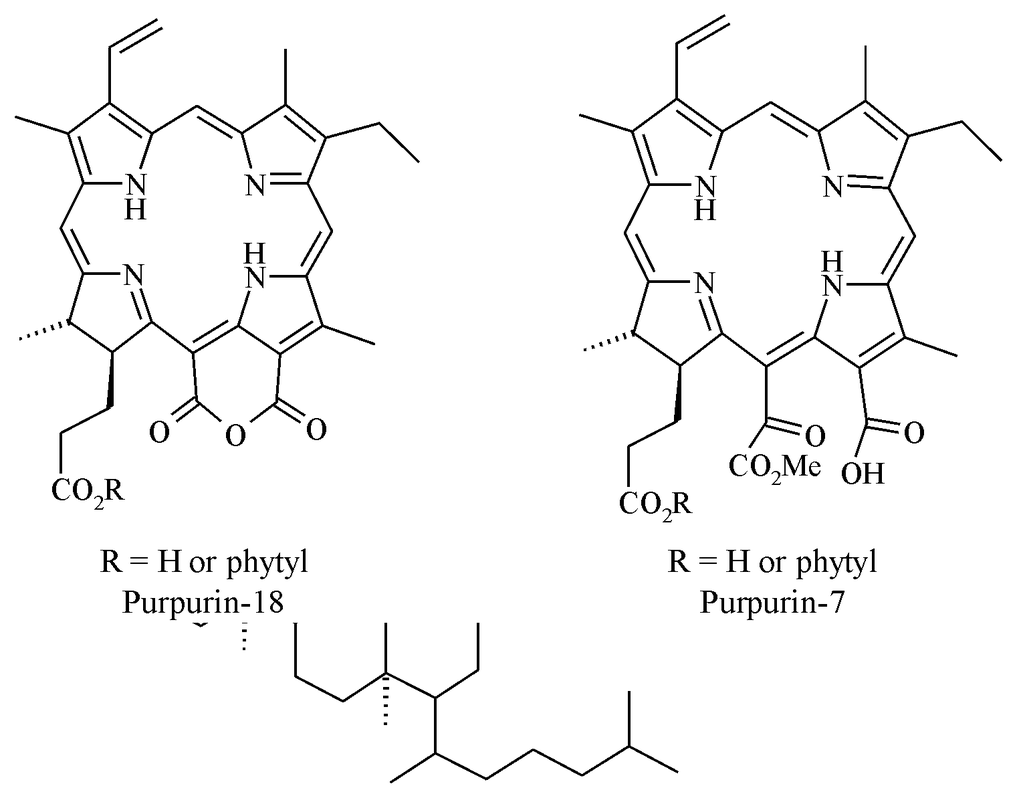
Figure 4: Structures of two chlorophyll oxidation products, purpurin-18 and purpurin-7.
|
Analysis of sedimentary pigment distributions can, therefore, allow particular environmental conditions to be inferred. As a result of the many transformation processes that affect chlorophyll pigments, their distributions in sediments are often complex, requiring rigorous methods of analysis.
Analysis of chlorophyll pigments
The combination of high performance liquid chromatography (HPLC) using a photodiode array detector (PDA) coupled with atmospheric pressure chemical ionisation (APCI) multistage mass spectrometry (MSn) permits comprehensive qualitative and quantitative analysis of pigment distributions as a linked sequence of events.
The components of the mixture are first separated by HPLC and the electronic absorption spectra of individual chromatographic peaks are recorded using the PDA. As a result of the highly conjugated nature of the pigments, the spectra reveal subtle differences in the nature of the pigment molecules and their transformation products. The eluent passes directly to the mass spectrometer where individual analytes are ionised by proton attachment, giving protonated molecules, [M+H]+, that reveal the molecular masses. Individual [M+H]+ ions are selected and subjected to collision-induced dissociation (CID) to produce product ions (the MS2 spectrum). Any one of the MS2 product ions can be selected for a further stage of CID to produce a new set of product ions (the MS3 spectrum). In this way a full mass spectral analysis can be performed within about five stages of CID. The losses observed during CID of chlorophyll and bacteriochlorophyll pigments and their derivatives are highly structure dependent, hence full structure assignment can often be made [7].
The combination of high performance liquid chromatography (HPLC) using a photodiode array detector (PDA) coupled with atmospheric pressure chemical ionisation (APCI) multistage mass spectrometry (MSn) permits comprehensive qualitative and quantitative analysis of pigment distributions as a linked sequence of events.
The components of the mixture are first separated by HPLC and the electronic absorption spectra of individual chromatographic peaks are recorded using the PDA. As a result of the highly conjugated nature of the pigments, the spectra reveal subtle differences in the nature of the pigment molecules and their transformation products. The eluent passes directly to the mass spectrometer where individual analytes are ionised by proton attachment, giving protonated molecules, [M+H]+, that reveal the molecular masses. Individual [M+H]+ ions are selected and subjected to collision-induced dissociation (CID) to produce product ions (the MS2 spectrum). Any one of the MS2 product ions can be selected for a further stage of CID to produce a new set of product ions (the MS3 spectrum). In this way a full mass spectral analysis can be performed within about five stages of CID. The losses observed during CID of chlorophyll and bacteriochlorophyll pigments and their derivatives are highly structure dependent, hence full structure assignment can often be made [7].
Porphyrin pigments in Antarctic lake sediments
The Larsemann Hills is an ice-free polar oasis in eastern Antarctica, located approximately midway between the eastern extremity of the Amery Ice Shelf and the southern boundary of the Vestfold Hills. At 50 km2, it is the second largest of only four major ice-free oases along East Antarctica’s 5000 km of coastline. More than 150 freshwater lakes are found in the hills, ranging from small ephemeral ponds to large water bodies such as Progress Lake (10 ha and 38 m deep).
The Larsemann Hills is an ice-free polar oasis in eastern Antarctica, located approximately midway between the eastern extremity of the Amery Ice Shelf and the southern boundary of the Vestfold Hills. At 50 km2, it is the second largest of only four major ice-free oases along East Antarctica’s 5000 km of coastline. More than 150 freshwater lakes are found in the hills, ranging from small ephemeral ponds to large water bodies such as Progress Lake (10 ha and 38 m deep).
Many low altitude coastal lakes (<15 m) also exist in this region and contain a valuable record of relative sea level changes from which regional ice-sheet history can be inferred. Kirisjes Pond is one such small freshwater lake in the Larsemann Hills region.
Pigment analysis of sediment from Kirisjes Pond. A sediment core from Kirisjes Pond, (12 Ha, 9 m depth, altitude 7 m), was obtained by Dr Dominic Hodgson (British Antarctic Survey) with the assistance of the Australian Antarctic Survey. The core was sectioned into 1 cm slices in the field and was kept frozen until analysis. For analysis the sediments were thawed, excess water was decanted and the pigments extracted by sonication in acetone. The sediment core from Kirisjes Pond showed three distinct pigment profiles (Figure 5).
Pigment analysis of sediment from Kirisjes Pond. A sediment core from Kirisjes Pond, (12 Ha, 9 m depth, altitude 7 m), was obtained by Dr Dominic Hodgson (British Antarctic Survey) with the assistance of the Australian Antarctic Survey. The core was sectioned into 1 cm slices in the field and was kept frozen until analysis. For analysis the sediments were thawed, excess water was decanted and the pigments extracted by sonication in acetone. The sediment core from Kirisjes Pond showed three distinct pigment profiles (Figure 5).
|
Kirisjes Pond -- pigments in the deepest section. The pigments in the deepest section (150 to 112 cm; c. 6525-11195 corr. Yrs BP) (corrected carbon-dated years before present) represent a phase during which freshwater conditions existed and supported a community of oxygen-evolving photosynthetic algae. This is indicated by the major pigments being chlorins derived from chlorophylls a and b, and polar carotenoids typical of oxygenic primary producers, and is supported by the presence of silicate fossils of freshwater diatoms (unicellular algae). Steryl chlorin esters are present in high relative abundance throughout this phase, suggesting an active population of zooplankton grazers was present in the lake. Trace amounts of purpurins indicate oxygenated conditions at the time of deposition.
Kirisjes Pond -- pigments in the middle section. The middle section of the core (94–110 cm; c. 6285–6525 corr. Yrs BP) shows a marked change in the primary producer population. The co-occurrence of bacteriochlorophylls c and d with chlorophyll a-derived pigments indicates the development of a population of photosynthetic bacteria accompanying the algal primary producers. |
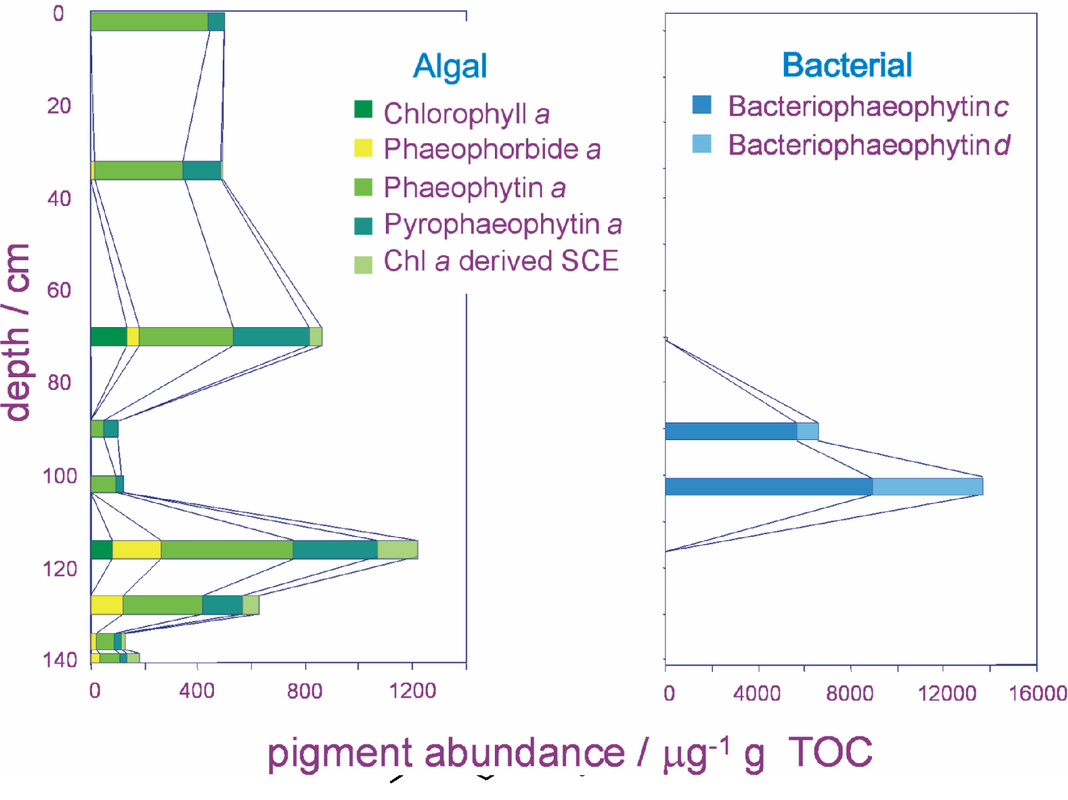
Figure 5: Depth profiles of chlorophyll (algal) and bacteriochlorophyll (bacterial) derivatives in the sediments of Kirisjes Pond, Antarctica
|
Such bacteria utilise H2S in photosynthesis in place of H2O and can only thrive under conditions where the water column is anoxic (excludes all oxygen). The presence of marine diatoms in the sediment indicates that a marine incursion occurred at the time this section was deposited. A subsequent transition zone (88–94 cm; c. 6205–6285 corr. Yrs BP) with a very high abundance of diatom cysts coincides with a shift in the distribution of bacteriophaeophytins c and d homologues to increased alkylation in the macrocycles. This may be related to the extent of development of the anaerobic community and the resultant increase in competition for the available light [5].
The high abundances of bacteriochlorophyll derivatives between 88 and 110 cm and very low abundance of chlorophyll a derivatives suggests this period represents development of a planktonic community dominated by anoxygenic photoautotrophs. In such environments grazing is severely restricted and pigment preservation is very good. Thus, aerobic productivity, as judged from the pigment concentrations, was suppressed during the time of the marine incursion.
Kirisjes Pond -- pigments from the uppermost section. The uppermost section of the core (surface--88 cm; c. present to 6205 Yrs BP) records a return to freshwater conditions with a dominance of oxygenic photosynthetic organisms. Freshwater diatoms are present and the major pigments are derived from chlorophylls a and b.
Differences between the two freshwater phases. A notable difference between the two freshwater phases is observed in the chlorophyll transformation products. While the total abundances of chlorophyll a and b components are approximately the same for both phases, significant variations in the distributions of the various transformation products are evident. Steryl chlorin esters are much less abundant in the upper phase, indicating a less well-developed community of grazers. The wider range of sterols present among the SCEs, with a greater relative contribution from sterols with 27 and 28 carbons, indicates a shift either in the primary producer community or the grazing community.
The non-SCE chlorophyll transformation products make a greater relative contribution to the total pigment distribution and the level of purpurins was higher than in the initial freshwater phase. Together, these features are suggestive of a greater proportion of the pigment distribution originating from dead and senescent cells than from deposition via faecal material as would be expected during times of greater grazing activity.
Kirisjes Pond -- pigments from the uppermost section. The uppermost section of the core (surface--88 cm; c. present to 6205 Yrs BP) records a return to freshwater conditions with a dominance of oxygenic photosynthetic organisms. Freshwater diatoms are present and the major pigments are derived from chlorophylls a and b.
Differences between the two freshwater phases. A notable difference between the two freshwater phases is observed in the chlorophyll transformation products. While the total abundances of chlorophyll a and b components are approximately the same for both phases, significant variations in the distributions of the various transformation products are evident. Steryl chlorin esters are much less abundant in the upper phase, indicating a less well-developed community of grazers. The wider range of sterols present among the SCEs, with a greater relative contribution from sterols with 27 and 28 carbons, indicates a shift either in the primary producer community or the grazing community.
The non-SCE chlorophyll transformation products make a greater relative contribution to the total pigment distribution and the level of purpurins was higher than in the initial freshwater phase. Together, these features are suggestive of a greater proportion of the pigment distribution originating from dead and senescent cells than from deposition via faecal material as would be expected during times of greater grazing activity.
|
Sea level changes and porphyrin distribution. The dramatic difference in the pigment profile in the middle section of the core can be explained as a consequence of change in relative sea level. As relative sea level increased the lake basin would have been inundated by marine waters. These waters would have contributed an increase in the sulfate levels in the lake, stimulating sulfate reducing bacteria in the lake bottom waters and/or sediment (Figure 6).
Sulfate reducing bacteria decompose organic matter by oxidation using sulfate as the source of oxidising power, thereby reducing the sulfate anions. The activity of the sulfate reducers would, in turn, have stimulated the photosynthetic bacterial community by supplying the H2S required for their photosynthetic activity. Increasing levels of H2S in the water column and increased consumption of oxygen through oxidation would have led to a stratified water column with algal photosynthesis being restricted to the oxygenated surface waters and with bacterial photosynthesis being prevalent within the anoxic deep waters of the lake. The absence of oxygen in the deep waters is reflected in the profiles of chlorophyll a oxidation products throughout the core.
Following a decrease in sea level around 6285 years ago, the lake reverted to a freshwater system dominated once again by algae. Notably, however, the activity of grazing organisms appears to have failed to reach previous levels and appears to have been eliminated during the more recent history of the lake. Chlorophyll oxidation products are detected in significant abundance only in the bottom and top sections of the core. Their presence only at very low levels in the middle section confirms the development of extensive water column anoxia during the period of connection to the ocean. |
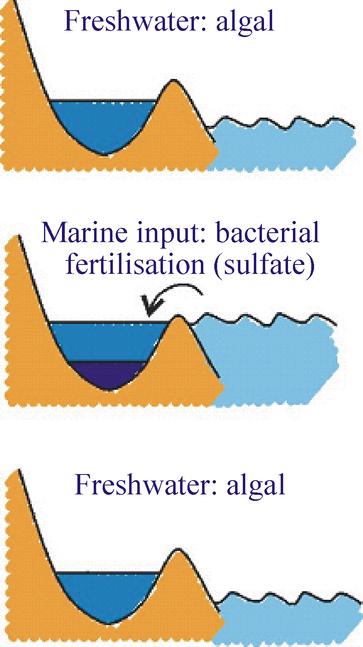
Figure 6: Diagram representing the three mainstages in the evolution
of Kirisjes Pondover the last 11,000 years (adapted from Squier, 2003,
reference 8)
|
Conclusion
Porphyrin pigment distribution found in cores from several other lakes in the region also reveal periods of connection with marine waters at similar times. Thus, the variation in relative sea level was a phenomenon that affected the whole of the region. The pigments are particularly useful markers of these environmental changes as they can be monitored readily, reflect all of the main groups of primary producers (unlike many fossil remains), and allow sediment records to be profiled at high stratigraphic resolution to generate records that have excellent time resolution.
Porphyrin pigment distribution found in cores from several other lakes in the region also reveal periods of connection with marine waters at similar times. Thus, the variation in relative sea level was a phenomenon that affected the whole of the region. The pigments are particularly useful markers of these environmental changes as they can be monitored readily, reflect all of the main groups of primary producers (unlike many fossil remains), and allow sediment records to be profiled at high stratigraphic resolution to generate records that have excellent time resolution.
Acknowledgement
The work described in this article formed part of the PhD studies of Angela Squier and was carried out in collaboration with Dr Dominic Hodgson of the British Antarctic Survey.
References
Dr BRENDAN KEELY,
Department of Chemistry,
University of York, Heslington,
York YO10 5DD
E-mail: [email protected]
Web link http://www.york.ac.uk/depts/chem/staff/bjk.html
This article is based on a presentation by Dr Keely at the ECG’s 2007 Distinguished Guest Lecture and Symposium ‘Environmental Chemistry in the Polar Regions’.
The work described in this article formed part of the PhD studies of Angela Squier and was carried out in collaboration with Dr Dominic Hodgson of the British Antarctic Survey.
References
- Vaughan, D. G.; Marshall, G. J.; Connolley, W. M.; Parkinson, C.; Mulvaney, R.; Hodgson, D. A.; King, J. C.; Pudsey, C. J.; Turner, J. (2003). Recent rapid regional climate warming on the Antarctic Peninsula, Climatic Change 60, 243.
- Knorr, G.; Lohmann, G. (2003). Southern Ocean origin for the resumption of Atlantic thermohaline circulation during deglaciation. Nature, 424, 532.
- Stocker, T. F. (2003). Global change: South dials north. Nature, 424, 496.
- Keely, B. J. (2006). In Chlorophylls and Bacteriochlorophylls: Biochemistry, Biophysics, Functions and Applications, (Advances in Photosynthesis and Respiration, Vol. 25, Series Editor: Govindjee), Grimm, B.; Porra, R. J.; Rüdiger, W.; Scheer, H. (Eds.), Springer-Verlag, Dordrecht, The Netherlands, pp. 535-561.
- Borrego, C. M.; Gerola, P. D.; Miller, M.; Cox, R. P. (1999). Light intensity effects on pigment composition and organisation in the green sulfur bacterium Chlorobium tepidum. Photosynthesis Research, 59, 159–166.
- Airs, R. L.; Atkinson, J. E.; Keely, B. J. (2001). Development and application of a high resolution liquid chromatographic method for the analysis of complex pigment distributions. Journal of Chromatography A. 917, 167-177.
- Wilson, M. A.; Hodgson, D. A.; Keely, B. J. (2005). Atmospheric pressure chemical ionisation liquid chromatography/multistage mass spectrometry for assignment of sedimentary bacteriochlorophyll derivatives. Rapid Communications in Mass Spectrometry, 19, 38-46.
- Squier A. H. (2003). PhD Thesis, University of York, UK.
Dr BRENDAN KEELY,
Department of Chemistry,
University of York, Heslington,
York YO10 5DD
E-mail: [email protected]
Web link http://www.york.ac.uk/depts/chem/staff/bjk.html
This article is based on a presentation by Dr Keely at the ECG’s 2007 Distinguished Guest Lecture and Symposium ‘Environmental Chemistry in the Polar Regions’.